- Prófase
- Metáfase
- Anáfase
- Telófase
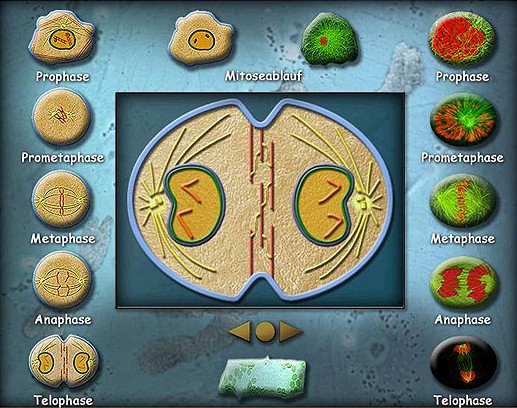
Na prófase:
- Os cromossomas, que foram duplicados durante a fase S da interfase, se condensam.
- O número de cromossomas varia de espécie para espécie, mas em humanos o número de cromossomas diplóides é 46.
- Os microtúbulos citoplasmáticos são desarranjados e a célula se prepara para a reorganização destes microtúbulos formando o fuso mitótico.
- O cromossoma mitótico consiste em dois cromatídios que estão conectadas por uma região denominada de centrómero.
- Na superfície deste centrómero existem dois cinetocoros, um deles está associado a cromátide.
- O outro cinetocoro está associado ao fuso mitótico e é através dele que resulta a movimentação cromossomal.
- Desorganização do envelope nuclear.
- A célula segue para a metáfase.
Durante a metáfase:
- Alguns dos microtúbulos que formam os aparatos do fuso se prendem aos cinetocoros formando o fuso mitótico.
- Os cromossomas iniciam uma série de movimentos que resultam num alinhamento de todos os cromossomas na região equatorial do fuso.
- É uma fase onde a célula se prepara para a anáfase.
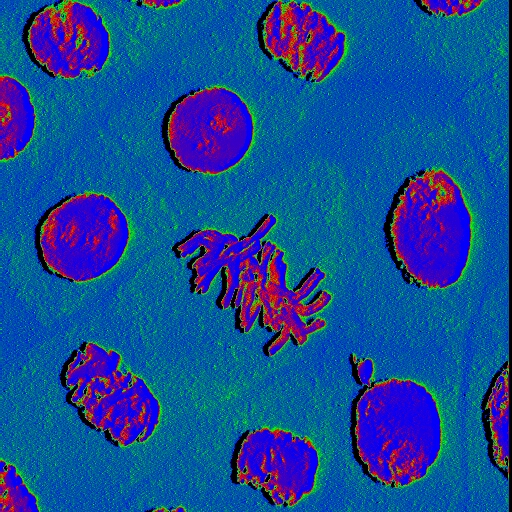
Anáfase:
- É o momento onde os cromatídios iniciam a migração para cada pólo da célula, em direcção aos centríolos, provocando a separação dos cromatídios irmãs.
- Acredita-se que a força que movimenta os cromatídios tem origem através da polimerização de proteínas dos microtúbulos (actina, miosina e tubulina).
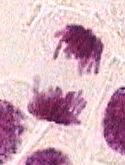
Término da anáfase e início da Telófase.
Durante a telófase:
- Separação completa dos cromatídios-irmãos para cada pólo da célula.
- Reconstituição do invólucro nuclear ao redor dos cromossomas.
- Descondensação dos cromossomas.
- Dissolução do aparato mitótico.
- Formação de uma constrição ao nível da zona equatorial da célula-mãe, que vai progredindo e termina por dividir o citoplasma e suas organelos em duas partes iguais.
- Neste ponto a célula termina a fase de divisão celular (Mitose) e entra na fase de replicação do DNA (Interfase) iniciando um novo ciclo.
Nesta fase, por microscopia, não visualizamos modificações tanto no citoplasma quanto no núcleo. As células porém estão em franca actividade, sintetizando os componentes que irão constituir as células filhas. O tempo de duração desta fase varia de célula para célula. É composta pela sucessão de três fases:
- G1 = Intervalo de tempo entre o final da mitose e o início da fase S
- S = Fase de Síntese de DNA
- G2 = Intervalo de tempo entre o final da fase S e o início da mitose
O tempo de duração da fase G1 é o principal factor para determinar o tempo da interfase.
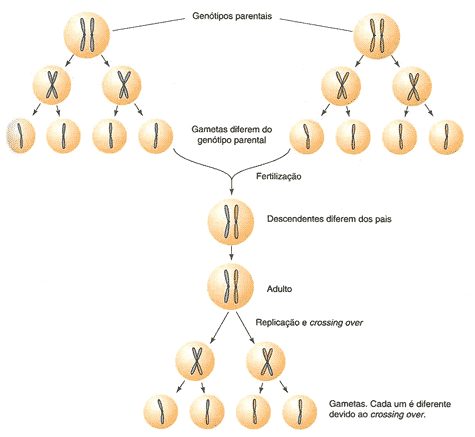
Um dos pressupostos fundamentais da biologia celular é o de que todas as células se originam a partir de células pré-existentes, à excepção do ovo ou zigoto que, nos seres vivos com reprodução sexuada, resulta da união de duas células reprodutivas (gâmetas), cada qual com metade da informação genética.
A mitose é um processo de divisão celular conservativa e tem quatro etapas definidas: Prófase, metáfase, anáfase e telófase. Denomina-se conservativa já que a partir de uma célula inicial, originam-se duas células idênticas, com igual composição genética (mesmo número e tipo de cromossomas), mantendo assim inalterada a composição e teor de DNA característico da espécie, excepto se ocorrer uma mutação. Este processo de divisão celular é comum a quase todos os seres vivos, dos animais e plantas multicelulares até aos organismos unicelulares, nos quais, muitas vezes, este é o principal ou único processo de reprodução – reprodução assexuada.
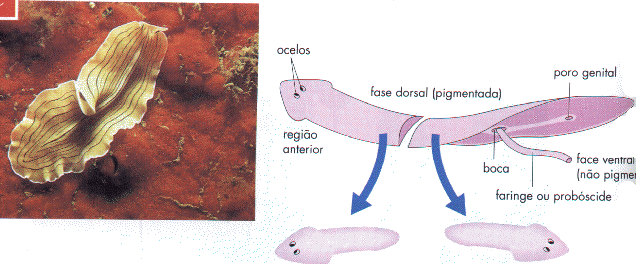
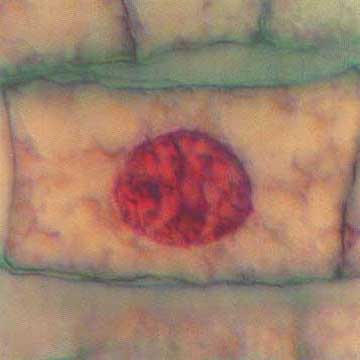
 Telofase numa célula vegetal
Telofase numa célula vegetal
Nas células vegetais, devido à presença de uma parede esquelética rígida, não é permitida a divisão por estrangulamento. Neste caso, vesículas derivadas do complexo de Golgi alinham-se na região equatorial e constituem o fragmoplasto. Estas vesículas fundem-se, para formar uma estrutura que é a membrana plasmática de cada célula filha. O conteúdo das vesículas vai originar a lamela mediana entre células filhas. Mais tarde, pela disposição de fibrilas de celulose, constituem-se as paredes esqueléticas. Estas paredes começam a formar-se da parte central para a periferia, até se ligarem à parede lateral da célula mãe. A parede celular não é contínua, deixando alguns poros, por onde os citoplasmas das células recém-formadas se continuam, constituindo os plasmodésmios, que testemunham a origem comum das duas células.
Outra diferença é o facto de nas plantas superiores, tal como acontece na cebola, não existem centríolos visíveis. As regiões correspondentes às duas zonas polares actuam como um centro organizador de microtúbulos que vão originar as fibrilas do fuso acromático.
posted by AS5MS @ 4:46 AM
2 comments
![]()